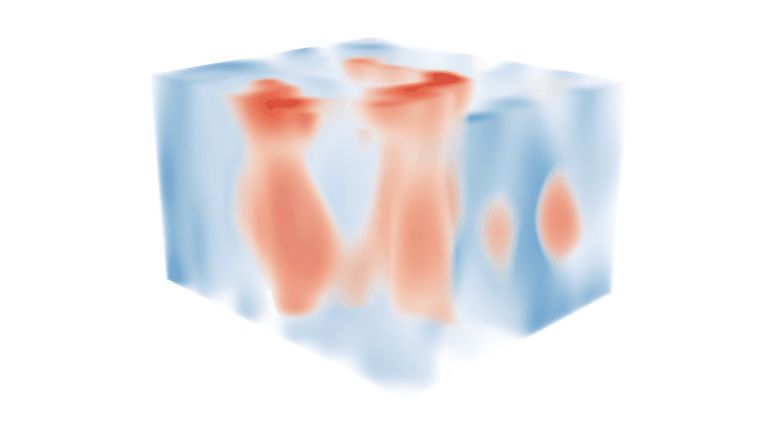
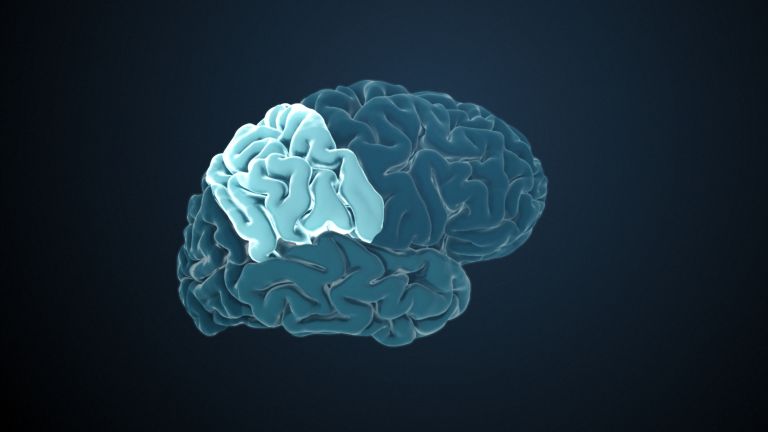
The Occipital Lobe
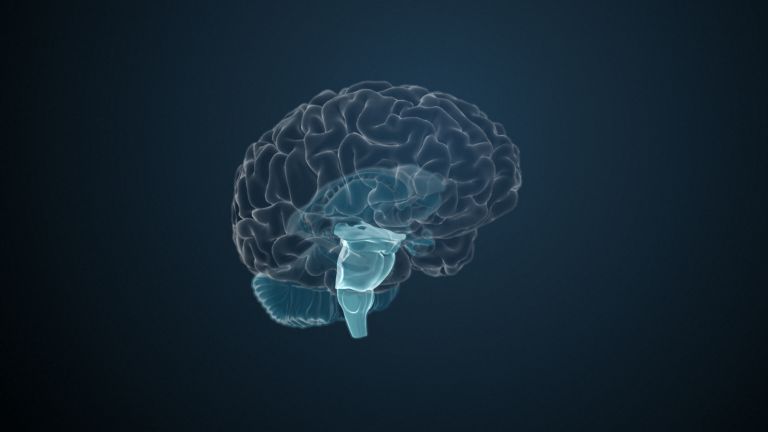
The occipital lobe is one of the most researched structures in the brain. This is partly due to its relatively manageable information inputs and partly due to its very clear, structured organization. Last but not least, the occipital lobe has a highly specialized area of responsibility: everything here revolves around vision.
Scientific support: Prof. Dr. Herbert Schwegler, Prof. Dr. Anne Albrecht
Published: 28.08.2025
Difficulty: intermediate
The Occipital lobe is responsible for processing visual impressions. In the primary visual cortex, every point in the field of vision is examined for lines and edges, movement and color. The fields of the secondary Visual cortex then use this information to develop more complex perceptions such as of space, color, movement, and even very specific objects. This complex processing impressively demonstrates how the brain receives and interprets signals from the outside world.
Occipital lobe
lobus occipitalis
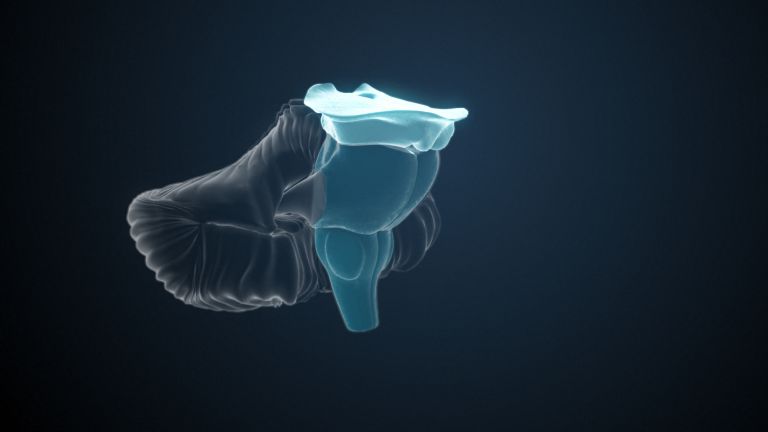
One of the four large lobes of the cerebral cortex. The occipital lobe lies above the cerebellum. It borders the parietal and temporal lobes at the front. The calcarine sulcus divides the occipital lobe into an upper and lower half, the cuneus and the lingual gyrus. Functionally, this area of the brain is responsible for the central processing of visual information – both the primary and secondary visual cortex are located in the occipital lobe.
Visual cortex
The visual cortex refers to the areas of the occipital lobe that are involved in processing visual information. These include the primary visual cortex and the associative visual cortices V1 to V5. According to Brodmann, the visual cortex comprises areas 17, 18, and 19.
The primary visual cortex
The Occipital lobe can be roughly divided into two areas: the primary visual cortex, or V1 for short, and the Visual association cortices V2 to V5. V1 corresponds to what is known as Brodmann area 17. Due to the special layering of the neurons in the cortex, this region has a stripe that is even visible to the naked eye, which is why the region is also called area striata. It is located largely on the medial, inward-facing side of the hemispheres at the occipital pole of the brain, thus forming the wall of the sulcus calcarinus.
V1 receives incoming nerve impulses, known as afferents, via optic radiation from the lateral geniculate Nucleus (CGL for short, part of the thalamus), whereby its 1.5 million fibers now face an impressive 200 million Cortex neurons. This may sound a little bureaucratic, but it is in fact a necessity for complex processing, as will become apparent.
The Primary visual cortex is retinotopically organized. This means that each point on the Retina corresponds to a specific small cortical area in V1, with neighborhoods remaining intact. Although the fovea, the point of sharpest vision on the retina, is only 1.5 millimeters in diameter, it occupies four-fifths of V1. This reflects the interconnection of cells in the retina, because in the fovea there is one Ganglion cell per photoreceptor – or to put it more simply: the “resolution” is particularly high here. Both ensure that we can process what we see in focus in the best possible way.
Occipital lobe
lobus occipitalis
One of the four large lobes of the cerebral cortex. The occipital lobe lies above the cerebellum. It borders the parietal and temporal lobes at the front. The calcarine sulcus divides the occipital lobe into an upper and lower half, the cuneus and the lingual gyrus. Functionally, this area of the brain is responsible for the central processing of visual information – both the primary and secondary visual cortex are located in the occipital lobe.
Visual association cortices
Area peristriata
The visual association cortices are the areas that are not involved in primary visual processing but play a major role in the interpretation and integration of visual information. They process, for example, shape, color, movement, or spatial orientation and forward information along the what pathway (ventral) for object recognition and the where/how pathway (dorsal) for spatial processing and action control.
\Brodmann area
As early as 1909, neuroanatomist Korbinian Brodmann divided the cerebral cortex into different areas. He did this based on histological criteria, distinguishing these areas according to their cellular structure. It later became apparent that these different structures of the cortex are often associated with different specializations.
lateral
A positional term – lateral means "towards the side." In relation to the nervous system, it refers to a direction at right angles to the neural axis, i.e., to the right or left.
Nucleus
In cell biology, the nucleus in a cell is the cell nucleus, which contains the chromosomes, among other things. In neuroanatomy, the nucleus in the nervous system refers to a collection of cell bodies – known as gray matter in the central nervous system and ganglia in the peripheral nervous system.
Cortex
cortex cerebri
Cortex refers to a collection of neurons, typically in the form of a thin surface. However, it usually refers to the cerebral cortex, the outermost layer of the cerebrum. It is 2.5 mm to 5 mm thick and rich in nerve cells. The cerebral cortex is heavily folded, comparable to a handkerchief in a cup. This creates numerous convolutions (gyri), fissures (fissurae), and sulci. Unfolded, the surface area of the cortex is approximately 1,800cm².
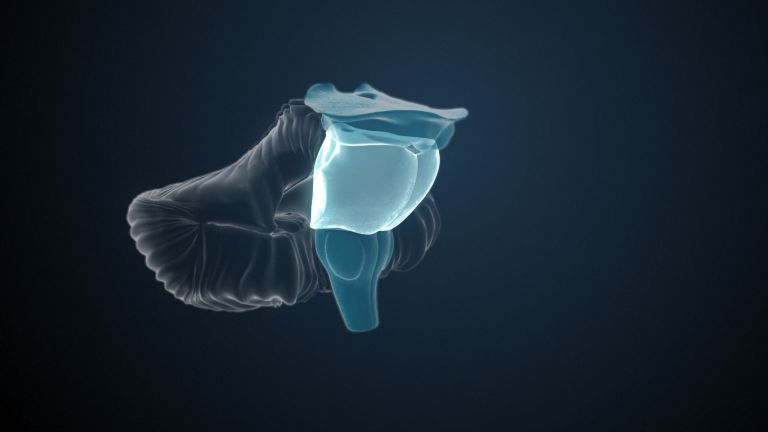
Primary visual cortex
area striata
The part of the occipital lobe whose primary inputs originate from the visual system. According to Brodmann, who originally divided the cerebral cortex into 52 areas in 1909, the primary visual cortex is area 17.
Retina
The retina is the inner layer of the eye covered with pigment epithelium. The retina is characterized by an inverse (reversed) arrangement: light must first pass through several layers before it hits the photoreceptors (cones and rods). The signals from the photoreceptors are transmitted via the optic nerve to the processing areas of the brain. The reason for the inverse arrangement is the evolutionary development of the retina, which is a protrusion of the brain.
The retina is approximately 0.2 to 0.5 mm thick.
Ganglion
Term for a cluster of nerve cell bodies in the peripheral nervous system. The term nerve node is often used because of its appearance. (Greek gágglion = knot-like)
Ganglion cell
The ganglion cell bundles the signals from the photoreceptors in the retina and transmits them via its axons (long, fiber-like extensions of a nerve cell. All of these axons together form the optic nerve.
Analysis at the cellular level
Like the entire isocortex, the Primary visual cortex is divided into six laminae, or layers, although it is significantly more complex than the rest of the Cortex. The individual layers differ in structure and function, but are strongly interconnected.
Of particular interest for sensory areas in general is layer IV, which receives the afferents of the sensory neurons – although the first processing steps already take place in the previous stations of the visual pathway, e.g., in the Retina and the CGL. Layer VI in the primary Visual cortex now receives this information in a rather complex interconnection pattern. It is particularly thick, so that it is subdivided into sublayers A, B, and C (the latter subdivided again into α and β).
Two examples of the network may suffice here: The cells of layer 4Cα mainly receive input from the magnocellular fibers of the GLC. These fibers carry information that primarily deals with the movement of objects and are in turn forwarded to layer 4B. There are also Parvocellular fibers, which are more object- and pattern-oriented. These mainly terminate in 4Cβ, and their cells in turn project into layers 2 and 3. This is already the beginning of the dorsal and the ventral streams of cortical visual processing.
In addition to this horizontal structure, there is also a vertical one, whose functional units were described as “columns” by Vernon Mountcastle (*1918) in the mid-1950s. Just as an aside: Nobel Prize winner David Hubel (*1926), who has done significant work in researching the visual system, would rather compare them to slices of toast.
Mountcastle's columns build on each other: several orientation columns – in which so-called “simple cells” detect the alignment of a line between 0 and 180 degrees –together form an Eye dominance column. Understanding their structure and distribution across the surface of the cortex proved to be quite difficult – comparable to trying to cut a lawn with nail scissors, as David Hubel later wrote. In fact, these “eye dominance columns” are not distributed alternately to the right and left, but overlap through horizontal and diagonal connections, becoming blurred at the boundaries. Today, a right and a left eye dominance column are combined to form a “hypercolumn,” each of which represents a small section of the outside world. This complex structure is one of the reasons for the enormous number of stimulus-processing cells in V1.
From above, the “blobs” descend into these orientation and eye dominance columns, which can only be detected using a special coloring technique. They are involved in the processing of color. David Hubel later explained the name “blobs” as follows: “We call them ‘blobs’ because the term is both vivid and unambiguous ... and because it seems to annoy our competitors.”
Blobs have a diameter of a quarter of a millimeter, in which five or six neurons can be derived. These neurons are often doubly complex in their behavior: for example, they react to red in the center with excitation and to central green with Inhibition. The opposite is true in the surrounding area. Blobs have no orientation specificity, unlike the area in between – the interblobs. However, the size of their area of responsibility can be calculated: approximately 200 different color tones in about 500 steps of Brightness and at least 26 levels of saturation result in over 2.5 million possibilities.
Primary visual cortex
area striata
The part of the occipital lobe whose primary inputs originate from the visual system. According to Brodmann, who originally divided the cerebral cortex into 52 areas in 1909, the primary visual cortex is area 17.
Cortex
cortex cerebri
Cortex refers to a collection of neurons, typically in the form of a thin surface. However, it usually refers to the cerebral cortex, the outermost layer of the cerebrum. It is 2.5 mm to 5 mm thick and rich in nerve cells. The cerebral cortex is heavily folded, comparable to a handkerchief in a cup. This creates numerous convolutions (gyri), fissures (fissurae), and sulci. Unfolded, the surface area of the cortex is approximately 1,800cm².
Retina
The retina is the inner layer of the eye covered with pigment epithelium. The retina is characterized by an inverse (reversed) arrangement: light must first pass through several layers before it hits the photoreceptors (cones and rods). The signals from the photoreceptors are transmitted via the optic nerve to the processing areas of the brain. The reason for the inverse arrangement is the evolutionary development of the retina, which is a protrusion of the brain.
The retina is approximately 0.2 to 0.5 mm thick.
Visual cortex
The visual cortex refers to the areas of the occipital lobe that are involved in processing visual information. These include the primary visual cortex and the associative visual cortices V1 to V5. According to Brodmann, the visual cortex comprises areas 17, 18, and 19.
Parvocellular
"Parvus" means "small." In the lateral geniculate nucleus, the switching station for visual stimuli in the thalamus, the outer four layers are called parvocellular because, unlike the magnocellular cell layers, they have small cell bodies. The parvocellular system transmits information for the perception of color and fine details.
dorsal
The positional term dorsal means "towards the back." In relation to the nervous system, it refers to a direction perpendicular to the neural axis, i.e., upwards towards the head or backwards.
In animals that do not walk upright, the term is simpler, as it always means toward the back. Due to the upright posture of humans, the brain bends in relation to the spinal cord, making dorsal mean "upward."
ventral
A positional term – ventral means "towards the abdomen." In relation to the nervous system, it refers to a direction perpendicular to the neural axis, i.e., downwards or forwards.
In animals (that do not walk upright), the term is simpler, as it always means toward the abdomen. Due to the upright posture of humans, the brain bends in relation to the spinal cord, making ventral mean "forward."
Eye
bulbus oculi
The eye is the sensory organ responsible for perceiving light stimuli – electromagnetic radiation within a specific frequency range. The light visible to humans lies in the range between 380 and 780 nanometers.
Inhibition
Neuronal inhibition describes the phenomenon whereby a sender neuron sends an impulse to a receiver neuron, causing the latter's activity to decrease. The most important inhibitory neurotransmitter is GABA.
Brightness
Brightness is one of the dimensions of visual perception: the perception of light intensity. It influences the size of the pupil.
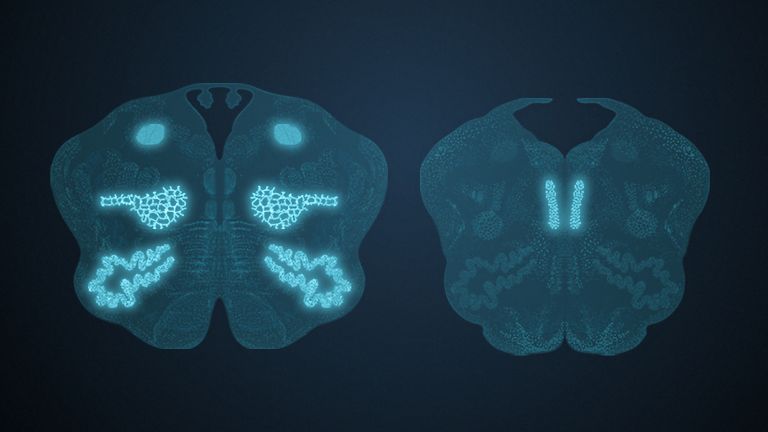
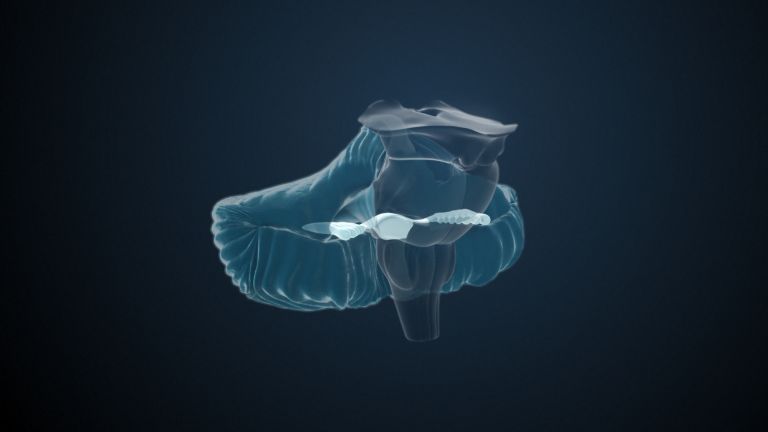
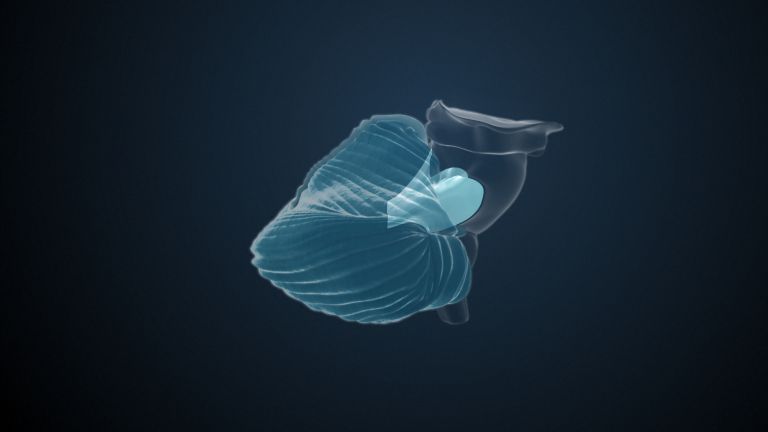
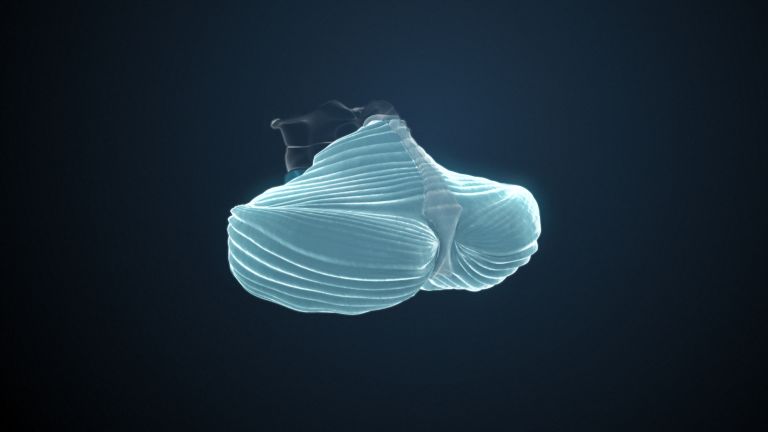

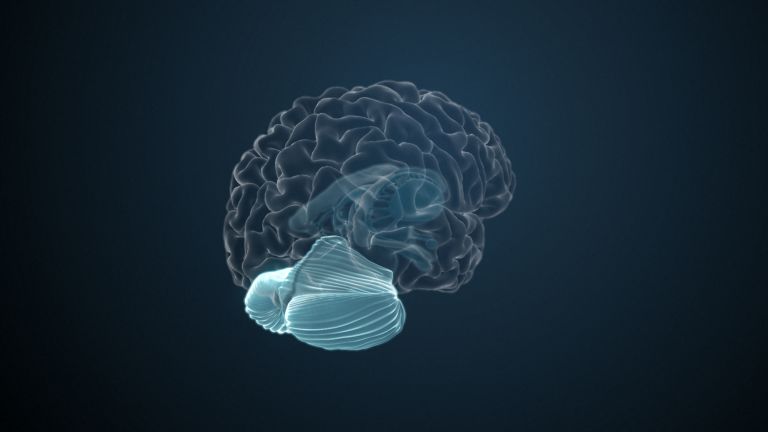
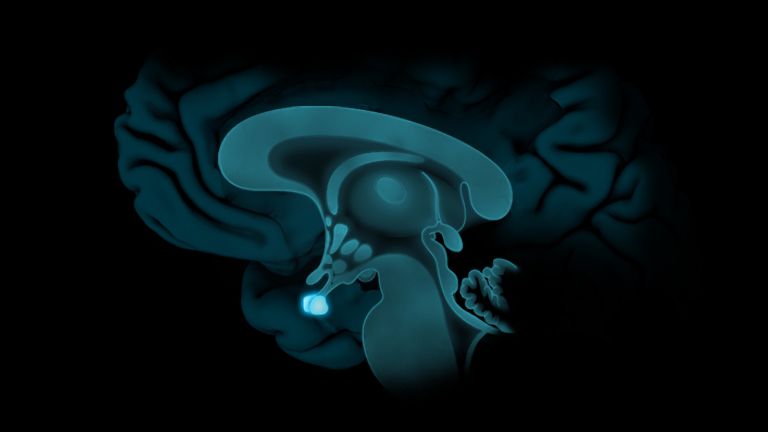

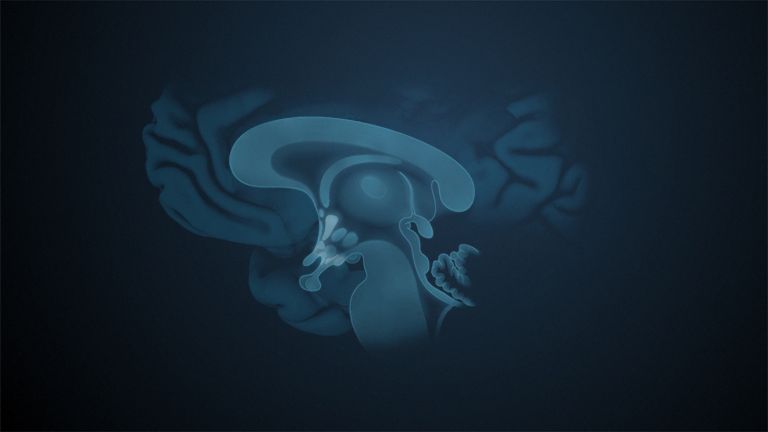

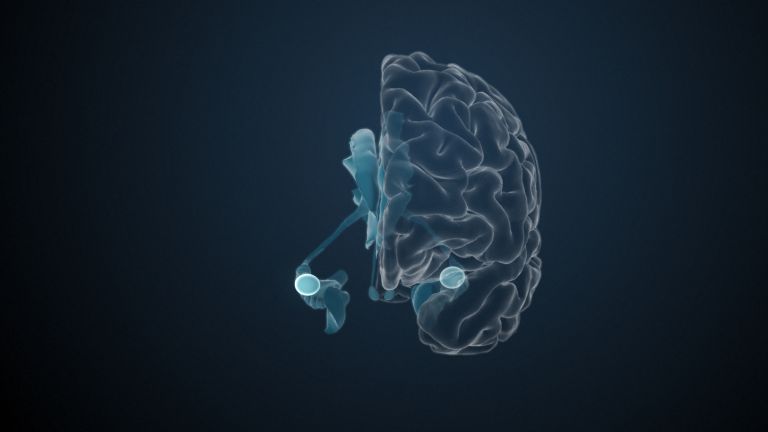
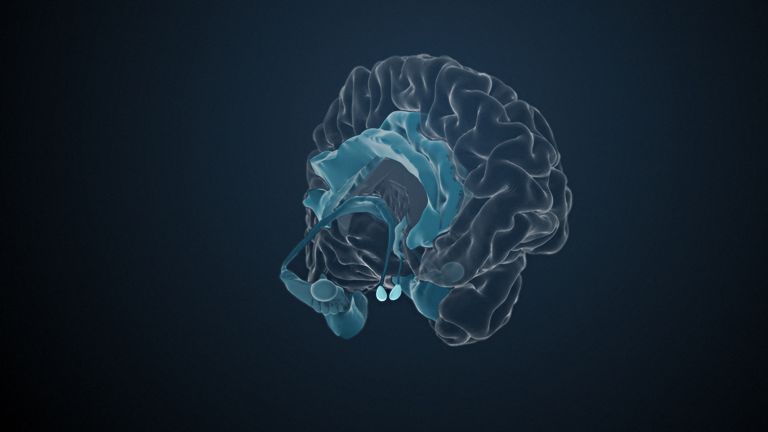
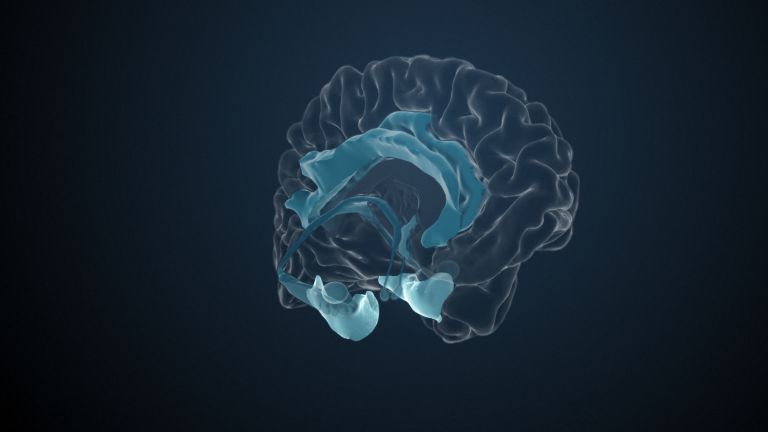
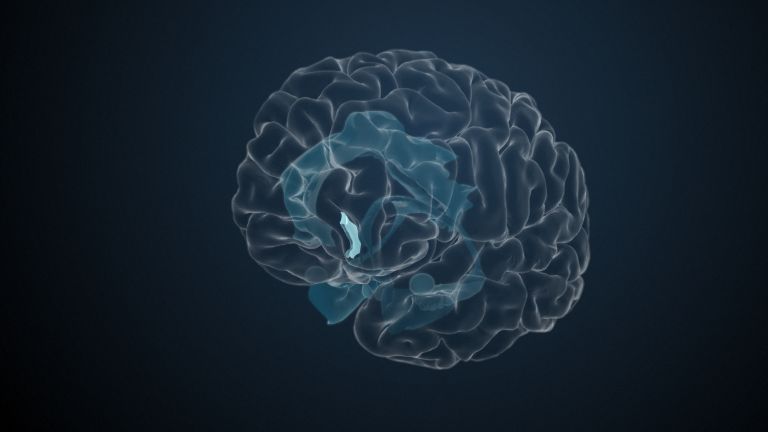
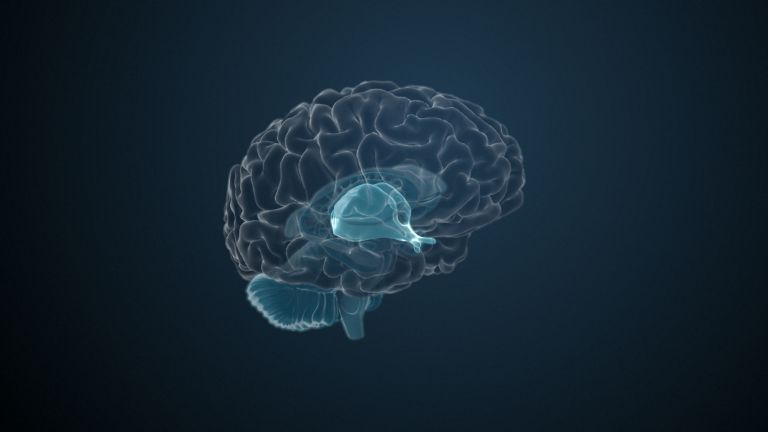
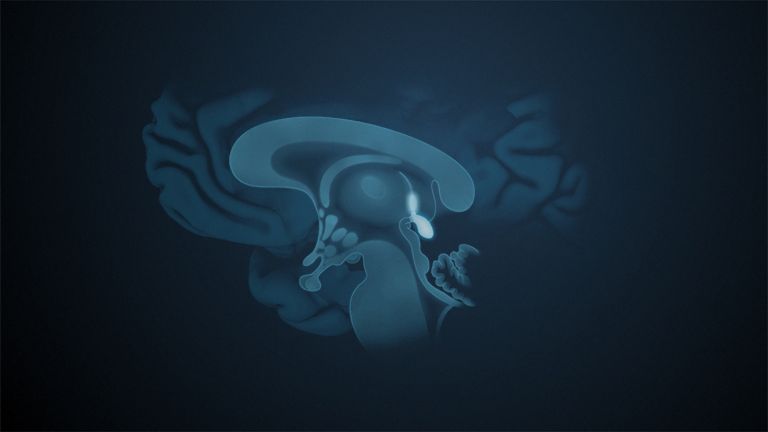
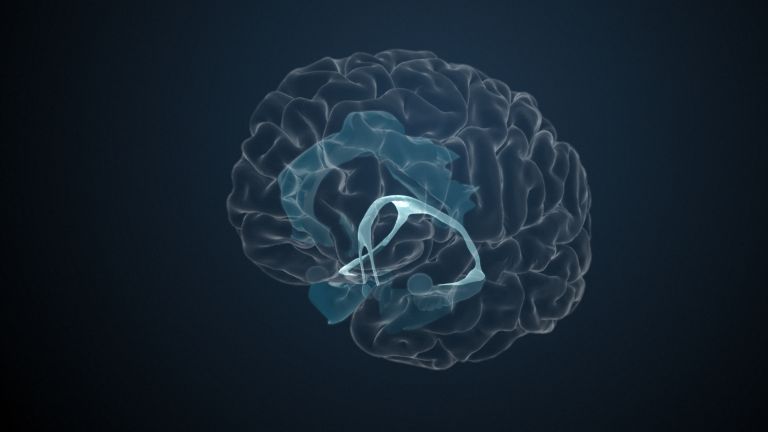
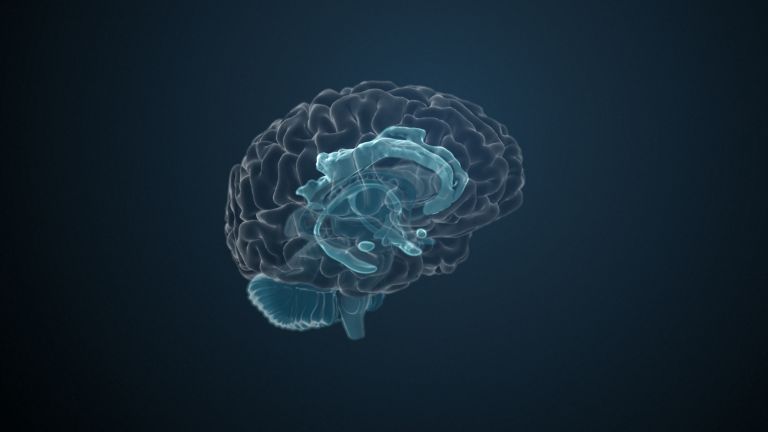
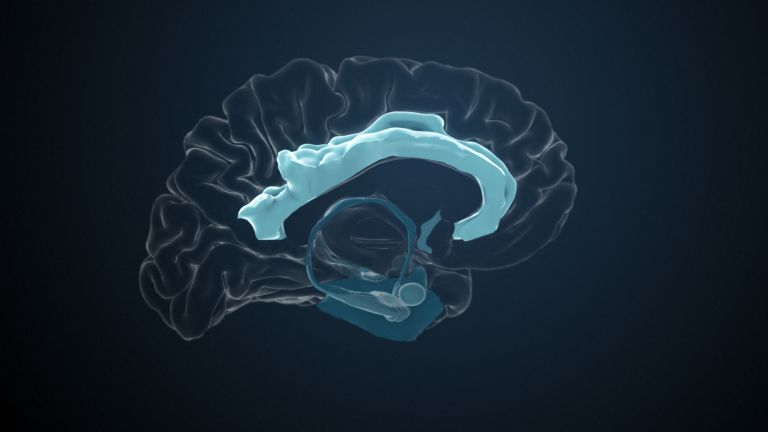
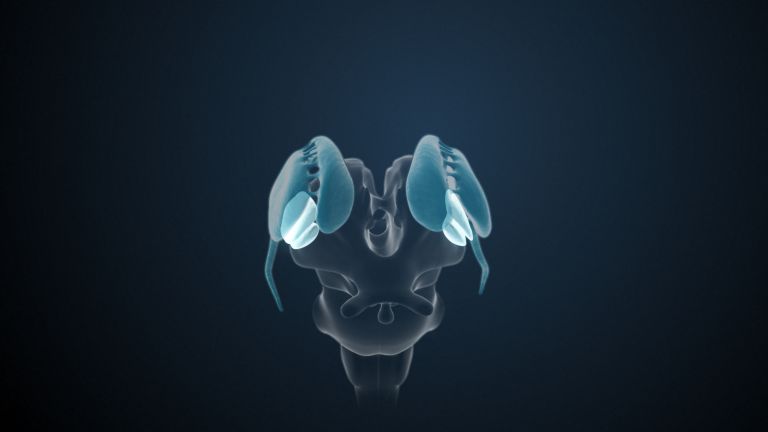
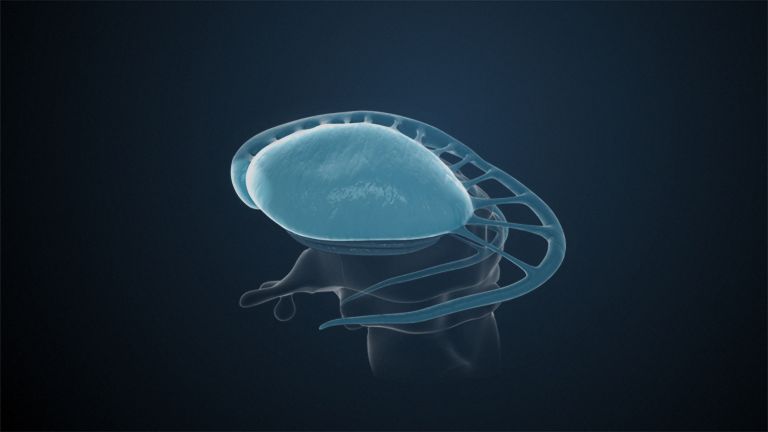
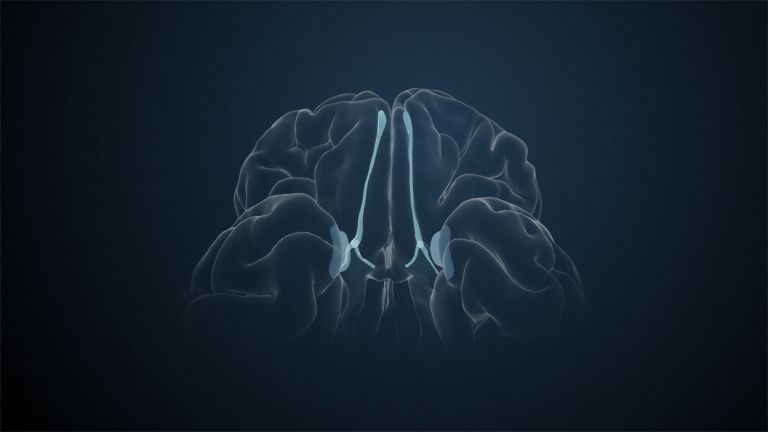
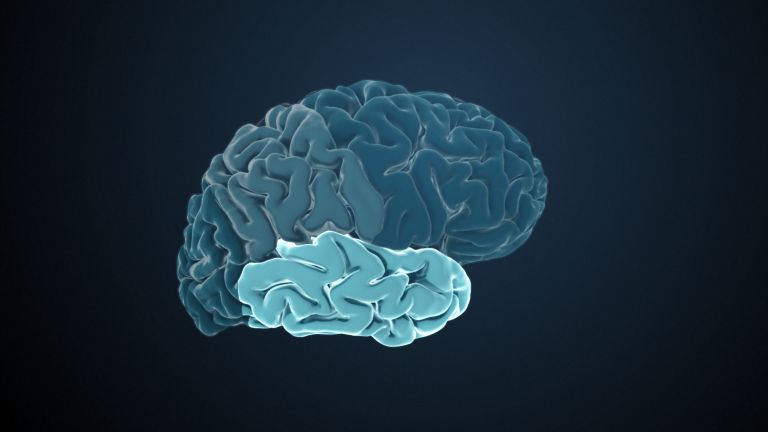
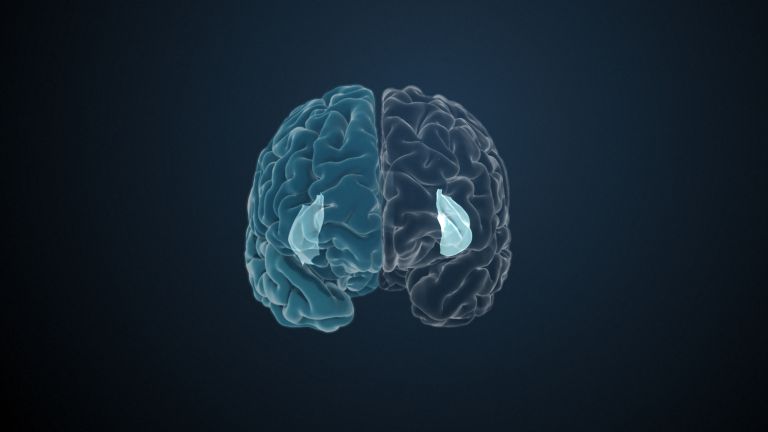
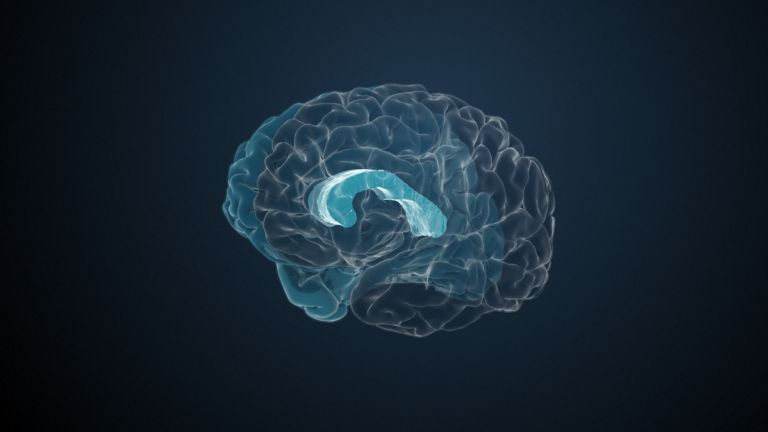
Visual association cortices
The Primary visual cortex is surrounded by several visual association areas. The classic secondary Visual cortex consists of V2 and V3 (according to Brodmann areas 18 and 19), whose inputs originate primarily from V1. The primary visual Cortex projects point by point and in an orderly manner onto the secondary visual cortex. Secondary processing spreads to other areas, while these higher processing areas simultaneously send feedback.
Overall, the magnocellular and Parvocellular pathways continue beyond the Occipital lobe – toward the Parietal lobe as the magnocellular dorsal stream, and toward the Temporal lobe as the parvocellular ventral stream. On both pathways, color, shape, movement, and space are increasingly refined, partly in areas with a very high degree of specialization. For example, there are areas for the Perception of faces or the distinction between familiar and unfamiliar objects.
Primary visual cortex
area striata
The part of the occipital lobe whose primary inputs originate from the visual system. According to Brodmann, who originally divided the cerebral cortex into 52 areas in 1909, the primary visual cortex is area 17.
Visual cortex
The visual cortex refers to the areas of the occipital lobe that are involved in processing visual information. These include the primary visual cortex and the associative visual cortices V1 to V5. According to Brodmann, the visual cortex comprises areas 17, 18, and 19.
Cortex
cortex cerebri
Cortex refers to a collection of neurons, typically in the form of a thin surface. However, it usually refers to the cerebral cortex, the outermost layer of the cerebrum. It is 2.5 mm to 5 mm thick and rich in nerve cells. The cerebral cortex is heavily folded, comparable to a handkerchief in a cup. This creates numerous convolutions (gyri), fissures (fissurae), and sulci. Unfolded, the surface area of the cortex is approximately 1,800cm².
Parvocellular
"Parvus" means "small." In the lateral geniculate nucleus, the switching station for visual stimuli in the thalamus, the outer four layers are called parvocellular because, unlike the magnocellular cell layers, they have small cell bodies. The parvocellular system transmits information for the perception of color and fine details.
Occipital lobe
lobus occipitalis
One of the four large lobes of the cerebral cortex. The occipital lobe lies above the cerebellum. It borders the parietal and temporal lobes at the front. The calcarine sulcus divides the occipital lobe into an upper and lower half, the cuneus and the lingual gyrus. Functionally, this area of the brain is responsible for the central processing of visual information – both the primary and secondary visual cortex are located in the occipital lobe.
Parietal lobe
Lobus parietalis
The parietal lobe is one of the four large lobes of the cerebral cortex. It is located behind the frontal lobe and above the occipital lobe. Somatosensory processes take place in its anterior region, while sensory information is integrated in its posterior region, enabling the handling of objects and spatial orientation. In addition, the parietal lobe is involved in attention, the recognition of body parts and objects, as well as linguistic and mathematical abilities.
dorsal
The positional term dorsal means "towards the back." In relation to the nervous system, it refers to a direction perpendicular to the neural axis, i.e., upwards towards the head or backwards.
In animals that do not walk upright, the term is simpler, as it always means toward the back. Due to the upright posture of humans, the brain bends in relation to the spinal cord, making dorsal mean "upward."
Temporal lobe
Lobus temporalis
The temporal lobe is one of the four lobes of the cerebrum and is located laterally (on the side) at the bottom. It contains important areas such as the auditory cortex and parts of Wernicke's area, as well as areas for higher visual processing; deep within it lies the medial temporal lobe with structures such as the hippocampus.
ventral
A positional term – ventral means "towards the abdomen." In relation to the nervous system, it refers to a direction perpendicular to the neural axis, i.e., downwards or forwards.
In animals (that do not walk upright), the term is simpler, as it always means toward the abdomen. Due to the upright posture of humans, the brain bends in relation to the spinal cord, making ventral mean "forward."
Perception
The term describes the complex process of gathering and processing information from stimuli in the environment and from the internal states of a living being. The brain combines the information, which is perceived partly consciously and partly unconsciously, into a subjectively meaningful overall impression. If the data it receives from the sensory organs is insufficient for this, it supplements it with empirical values. This can lead to misinterpretations and explains why we succumb to optical illusions or fall for magic tricks.
Deficits
Lesions in the Primary visual cortex prevent the processing of corresponding impulses at the affected site. This results in deficits in the visual field – in the worst case, complete destruction of V1 results in what is known as Cortical blindness although the Retina and Visual pathway are intact, the patient is completely blind.
Deficits in the secondary Visual cortex do not lead to visual field defects. Rather, patients are no longer able to classify or recognize what they see. This can affect colors, shapes, or even faces. In this case, the condition is referred to as Visual agnosia Synesthesia and visual hallucinations also appear to be caused by disorders of the secondary visual cortex.
Primary visual cortex
area striata
The part of the occipital lobe whose primary inputs originate from the visual system. According to Brodmann, who originally divided the cerebral cortex into 52 areas in 1909, the primary visual cortex is area 17.
Cortical blindness
When the primary visual cortex is destroyed, those affected are effectively blind, even though their eyes are undamaged. However, they are often unconsciously able to grasp and catch objects and even name them when asked to guess. This effect is known as blind sight.
Retina
The retina is the inner layer of the eye covered with pigment epithelium. The retina is characterized by an inverse (reversed) arrangement: light must first pass through several layers before it hits the photoreceptors (cones and rods). The signals from the photoreceptors are transmitted via the optic nerve to the processing areas of the brain. The reason for the inverse arrangement is the evolutionary development of the retina, which is a protrusion of the brain.
The retina is approximately 0.2 to 0.5 mm thick.
Visual pathway
The visual pathway refers to the network of nerve cells involved in visual perception. In mammals, it runs from the retinal ganglion cells in the eye – as the optic nerve to the optic chiasm, then as the visual tract – via the only switching point in the lateral geniculate nucleus to the primary visual cortex.
Visual cortex
The visual cortex refers to the areas of the occipital lobe that are involved in processing visual information. These include the primary visual cortex and the associative visual cortices V1 to V5. According to Brodmann, the visual cortex comprises areas 17, 18, and 19.
Visual agnosia
Agnosia can be translated as "not recognizing." It refers to deficits in visual perception without blindness. The term was coined by Sigmund Freud. Examples of visual agnosia include prosopagnosia (face agnosia) and object agnosia.
Synesthesia
Synesthesia is the linking of two or more sensory perceptions to form a subjective sensation. For example, a synesthete might always associate the number seven with the color red. Synesthesia appears to have a hereditary component, but it can also be caused by illness (e.g., schizophrenia) or drugs (e.g., hallucinogens).
Cortex
cortex cerebri
Cortex refers to a collection of neurons, typically in the form of a thin surface. However, it usually refers to the cerebral cortex, the outermost layer of the cerebrum. It is 2.5 mm to 5 mm thick and rich in nerve cells. The cerebral cortex is heavily folded, comparable to a handkerchief in a cup. This creates numerous convolutions (gyri), fissures (fissurae), and sulci. Unfolded, the surface area of the cortex is approximately 1,800cm².
First published on September 23, 2011
Last updated on August 28, 2025